Afdholiatus Syafaah, M.Sc.

Afdholiatus Syafaah, M.Sc.
Peneliti Pemuliaan Tanaman
- telp
- youtube.com

Pendidikan
Minat Riset
Biografi Singkat
Sigit memperoleh gelar Sarjana (Biologi) dari Fakultas Biologi, Universitas Gadjah Mada pada tahun 2004. Gelar Master of Science diperoleh dari University Putra Malaysia pada tahun 2013. Pada tahun 2017 memperoleh jenjang fungsional Peneliti Muda Bidang Genetika dan Pemuliaan Tanaman.

Pendidikan
Minat Riset
Biografi Singkat
Fetrina memperoleh gelar Sarjana Sains dari Universitas Andalas pada tahun 1999. Gelar Magister Sains diperoleh pada tahun 2004 dan Doktor pada tahun 2016 dari Institute Pertanian Bogor. Tahun 2018 memperoleh jenjang fungsional Peneliti Madya. Tahun 2019 - 2020 menjabat sebagai Koordinator Penelitian Balai Penelitian Sembawa, dilanjutkan dengan Kepala Sub Operasional Penelitian Pusat Penelitian Karet. Sejak 2019 sampai saat ini sebagai Ketua Kelompok Penelian Pemuliaan Pusat Penelitian Karet.
Fetrina aktif melakukan kegiatan penelitian yang mendukung program pemuliaan tanaman karet di Indonesia. Dalam pengembangan kerja sama International, Fetrina berpartisipasi aktif dalam kegiatan Breeding Group of IRRDB (International Rubber Research Development Board) sebagai Country representative dari Indonesia. Selain itu Fetrina juga berkesempatan berkolaborasi dengan Cirad, Perancis dan Agromillora, Spanyol mengembangkan bahan tanam karet asal kultur jaringan pada tahun 2017-2021. Kolaborasi dengan Cirad juga dilakukan terkait dengan Genome and Genetic Analysis of Tapping Panel Drynes pada tahun 2016-2020 dilanjutkan dengan co-coordinator kegiatan kerjasama penelitian Rubis Project konsorsium Pusat Penelitian Karet, Cirad dan Universitas Gajah Mada yang mendapat pendanaan dari Agropolis Foundation, Perancis. Pada 2018 juga melakukan kolaborasi penelitian dengan Riken, Jepang terkait dengan Research on breeding of natural rubber yang dilanjutkan dengan project Satrep pada tahun 2021. Pada saat yang sama juga dilakukan pengembangan Natural rubber properties melalui kerja sama dengan Riken dan Yokohama company, Jepang.

Pertumbuhan dan produktivitas tanaman karet (Hevea brasiliensis) ditentukan oleh faktor genotipe (G), faktor lingkungan (E), dan interaksi genotipe x lingkungan (GxE). Kondisi lingkungan yang sangat berpengaruh terhadap produksi karet salah satunya adalah kondisi curah hujan. Tanaman karet memiliki sifat menggugurkan daun pada setiap musim kemarau, gugur daun terjadi seiring adanya perubahan pola curah hujan bulanan. Kondisi curah hujan rendah mengakibatkan tanaman karet menggugurkan daun secara alami sebagai respon terhadap cekaman kekeringan yang terjadi pada bulan-bulan kering setiap tahunnya. Setiap klon tentunya akan memiliki respon yang berbeda terhadap perubahan jumlah curah hujan yang terjadi pada setiap bulannya. Gugur daun fisiologi yang terjadi pada tanaman karet merupakan respon tanaman untuk mencegah transpirasi berlebih pada saat terjadinya cekaman kekeringan. Secara genetik tentunya masing-masing klon memiliki mekanisme adaptasi yang berbeda pada saat terjadinya perubahan lingkungan.
Dinamika gugur daun pada tanaman karet terdiri dari lima fase yaitu fase 1 yang ditandai dengan muncul tanda-tanda daun menguning sampai daun kuning sebagian, fase 2 yang ditandai dengan kondisi daun kuning menyeluruh dan sebagian lagi sudah gugur, fase 3 ditandai dengan semua daun gugur dan muncul kuncup daun berwarna cokelat, fase 4 ditandai dengan daun mulai berwarna hijau muda, dan fase 5 ditandai dengan kondisi daun berwarna hijau tua.
Fase perdaunan pada tanaman karet
Dari hasil penelitian yang telah dilakukan menunjukkan bahwa dinamika fase gugur daun memiliki pengaruh nyata terhadap hasil lateks.
Pengaruh fase daun terhadap hasil lateks dari beberapa genotype karet
Jika ditinjau dari dinamika fase gugur daun menunjukkan bahwa hasil lateks (g/p/s) paling tinggi terdapat pada fase 5, sedangkan paling rendah terjadi pada kondisi daun fase 3 dan 4. Hasil lateks pada kondisi daun fase 1 tidak berbeda dengan fase 2. Fluktuasi hasil lateks sangat dipengaruhi oleh kondisi daun tanaman. Kondisi perdaunan karet memiliki hubungan yang erat kaitannya dengan distribusi curah hujan. Turunnya kadar air tanah pada saat musim kemarau akan mempengaruhi penyerapan air dan unsur hara tanaman yang selanjutnya akan mempengaruhi metabolisme dan pertumbuhan tanaman. Salah satu fungsi utama air bagi tanaman adalah mempertahankan turgiditas sel dan jaringan tanaman yang penting bagi kelangsungan aktivitas sel dalam pembelahan dan pemanjangan sel. Pengaruh langsung yang terjadi akibat kekurangan air berkepanjangan yaitu berkurangnya laju pertumbuhan, sehingga ukuran tanaman dan hasil lateks rendah dibandingkan saat tanaman dalam kondisi normal.
Tanaman yang memiliki kecukupan air akan lebih efektif dalam kegiatan fotosintesis untuk menghasilkan asimilat. Kapasitas fotosintesis tanaman karet menurun saat gugur daun, sehingga hasil lateks pada klon karet umumnya juga menurun. Penurunan hasil lateks terjadi secara nyata pada saat pembentukan kuncup daun dan daun muda yaitu pada fase 3 dan 4. Hasil lateks yang rendah pada fase tersebut diduga disebabkan karbohidrat yang dihasilkan pada saat fotosintesis yang terdapat pada tanaman lebih diutamakan untuk pertumbuhan tanaman dibandingkan untuk mensintesis partikel karet.
Referensi:
Oktavia, F., dan Lasminingsih, M. (2010). Pengaruh kondisi daun tanaman karet terhadap keragaman hasil sadap beberapa klon seri IRR. J. Penel. Karet. 29 (2): 32-40.
Priyadarshan, P.M., Sasikumar, S., and Concalves, D.(2001). Phenological changes in Hevea brasiliensis under differential geo climates. The Planter. 77: 447-481.
Siregar, T.H.S., Tohari, Hartiko, H., dan Karyudi. (2007). Dinamika perontokan dan pohon karet dan hasil lateks: I. Jumlah daun rontok dan hasil lateks. J. Penel. Karet, 25(1): 45-58.
Thomas dan Boerhendhy, I. (1988). Hubungan neraca air tanah dengan produksi karet klon GT 1 dan PR 261. Bull Perkebunan Rakyat. 4(1): 15-18.
1905
Penyakit Gugur Daun Colletotrichum
(Colletotrichum gloeosporioides dan C. acutatum)
Pertama kali dilaporkan di Sri Lanka dan negara lainnya (Jayasinghe et al. 1997). Kerugian produksi lateks akibat penyakit ini sebesar 7% sampai 45% tergantung dari intensitas serangan patogen (Saha et al., 2002; Guixiu, 2014).
1918
Penyakit Gugur Daun Colletotrichum
Pertama kali dilaporkan di Indonesia. Gejala pada daun muda adalah daun mengeriput, menggulung, ujung daun mati, dan gugur. Gejala pada daun tua terdapat bercak kecil berwarna hitam, berlubang, dan bagian ujung mati. Serangan berat menyebabkan gugur daun dan tajuk meranggas (Shufen et al., 1999).
Penyakit Hawar Daun Amerika Selatan
(Microcyclus ulei)
Pertama kali dilaporkan di Suriname, Amerika Selatan (Lieberei, 2007). Di Indonesia cendawan M. ulei merupakan OPTK A1 yang berarti cendawan ini tidak terdapat di wilayah Indonesia dan dicegah masuknya ke dalam wilayah Indonesia (Peraturan Menteri Pertanian RI Nomor 51 Tahun 2015).
1938
Penyakit Gugur Daun Oidium
(Oidium heveae)
Pertama kali dilaporkan di Indonesia. Patogen menyerang daun karet muda, kuncup bunga, dan jaringan muda lainnya sehingga mengurangi hasil karet hingga 45% (Liyanage et al., 2016).
1958
Penyakit Gugur Daun Corynespora
(Corynespora cassiicola)
Pertama kali ditemukan di India, kemudian pada tahun 1960 ditemukan di Malaysia dan 1966 di Nigeria (Jayasinghe & Fernando, 2011).
1980
Penyakit Gugur daun Corynespora
Pertama kali dilaporkan di Indonesia. Patogen menyerang daun yang masih muda atau berwarna kecokelatan. Gejala pada daun karet terdapat bercak seperti sirip ikan dan warna daun menjadi kuning atau cokelat kemudian gugur (Situmorang et al., 2004).
1987
Penyakit Hawar daun Fusicoccum
(Neofusicoccum ribis)
Pertama dilaporkan di Johor, Malaysia (Radziah & Chee, 1989), kemudian pada tahun 2003 dilaporkan kembali di Perak, Malaysia (Mahyudin & Zamri, 2018). Cendawan N. ribis merupakan OPTK A2 yang penyebarannya terbatas di wilayah Sumatera, Jawa, dan Kalimantan (Peraturan Menteri Pertanian RI Nomor 51 Tahun 2015). Gejala khas pada daunmunculnya zona kecoklatan konsentris pada permukaan atas daun (Nyaka et al., 2012).
2013
Penyakit Gugur Daun Pestalotiopsis
(Pestalotiopsis microspora)
Dilaporkan terdapat di Cameroon (Nyaka et al., 2017). Gejala penyakit berupa bercak daun berbentuk bulat yang terdapat pada daun berwarna hijau sampai hijau tua. Daun yang terserang patogen dapat mengalami perubahan warna menjadi kuning atau oranye.
2017
Penyakit Gugur daun Pestalotiopsis
Penyakit ditemukan di beberapa wilayah di Indonesia. Penyakit juga dilaporkan berkembang luas di beberapa negara Asia Tenggara seperti Malaysia, India, Thailand, dan Sri Lanka. Adanya kejadian penyakit tersebut menyebabkan terjadinya penurunan produksi lateks sampai 46% (Kusdiana et al., 2020).
Alchemi Putri Juliantika Kusdiana, M.Si.
Peneliti Proteksi Pusat Penelitian Karet
Sumber:
Jayasinghe CK, Fernando THPS, Priyanka UMS. 1997. Colletotrichum acutatum is the main cause of Colletotrichum leaf disease of rubber in Sri Lanka. Mycopathologia. 137:53–56. doi: 10.1023/A:1006850119146.
Jayasinghe CK, Fernando THPS. 2011. Corynespora Leaf Fall of Hevea Rubber the Most Threatening Leaf Disease in Asia & African Continents. Malaysia (MY): CFC & IRRDB.
Kusdiana APJ, Sinaga MS, Tondok ET. 2020. Diagnosis Penyakit Gugur Daun Karet (Hevea brasiliensis Muell. Arg.). Jurnal Penelitian Karet. 38(2). doi: 10.22302/ppk.jpk.v2i38.728.
Lieberei, R. 2007. South American Leaf Blight of the Rubber Tree (Hevea spp.): New Steps in Plant Domestication using Physiological Features and Molecular Markers. Ann Bot. 100(6): 1125–1142. doi: 10.1093/aob/mcm133.
Liyanage KK, Khan S, Mortimer PE, Hyde KD, Xu J, Brooks S, Ming Z. 2016. Powdery mildew disease of rubber tree. Forest Pathology. 46(2):90–103. doi: 10.1111/efp.12271.
Mahyudin MM, Zamri AMA. 2018. Strategies to minimize the incidence of Fusicoccum leaf blight. International Plant Protection Workshop 2018. Palembang: IRRI & IRRDB.
Nyaka NAIC, Abidin MAZ, Wong MY, Murnita MM. 2012. Cultural and morphological characterisations of Fusicoccum sp., the causal agent of rubber (Hevea brasiliensis) leaf blight in Malaysia. Journal of Rubber Research. 15(1):64-79.
Nyaka NAIC, Owona PAN, Oumar D, Ntsomboh GN, Njonje SW, Ehabe EE. 2017. Characterization of Pestalotiopsis microspora, causal agent of leaf blight on rubber (Hevea brasiliensis) in Cameroon. Proceedings International Rubber Conference 2017. Bogor: IRRDB & IRRI.
Radziah NZ, Chee KH. 1989. A new foliar disease of rubber. Plant Pathology. 38:293-296. doi: 10.1111/j.1365-3059.1989.tb02147.x.
Shufen F, Gang G, Fucong Z. 1999. General situation of anthracnose of rubber trees and its researches in China. Proceedings of IRRDB Symposium 1999. China:Hainan Publishing House.
Situmorang A, Sinaga MS, Suseno R, Hidayat SH, Siswanto, Darussamin A. 2004. Status dan manajemen pengendalian penyakit gugur daun Corynespora di perkebunan karet. Prosiding Pertemuan Teknis. Palembang: Pusat Penelitian Karet.


Penyadapan karet menggunakan irisan ganda (double cut/DC) sudah lebih dari 1 dasawarsa diterapkan di perkebunan karet, yaitu sejak dikenalkan sistem sadap berdasarkan perbedaan metabolisme lateksnya, atau dikenal dengan istilah sistem sadap berdasarkan tipologi klon.
Penyadapan DC dilakukan pada klon yang metabolisme lateksnya tergolong sedang dan rendah, seperti GT 1, BPM 1, BPM 24, BPM 107, BPM 109, PR 255, PR 261, PR 300, PR 303, PB 217, PB 330, RRIC 100, RRIC 102, RRIC 110, RRIM 717, IRR 9, AVROS 2037, TM 2, TM 6, TM 8, dan TM 9. Sistem sadap DC diterapkan di panel BI dan H0, umumnya dimulai sejak umur TM ke-11. Penyadapan pada klon tersebut dilakukan dengan cara mengiris panel kulit pulihan (BI) S/2 dan dikombinasi panel atas (H0) S/4U + etefon. Penambahan irisan ¼SU + etefon di panel atas (H0) melalui penyadapan DC terbukti dapat mengoptimalkan perolehan produksi ketika panel bawah (BI) melemah.
Penerapan sistem sadap DC seringkali di luar ekspektasi. Hal yang jelas terlihat adalah konsumsi kulit semenjak disadap DC terkesan semakin boros dan tren produksi menurun. Padahal, sebenarnya hal tersebut terjadi akibat kurang disiplin penjagaan mutu sadap dan lemahnya kontrol sadap, sehingga mutu sadap jelek. Mutu sadap jelek utamanya ditandai dengan irisan tebal (>2,5 mm/sadap), panjang irisan panel atas (H0) > S/4U dan banyak luka kayu. Dampak jangka panjang yang timbul adalah panel habis sebelum waktunya. Hasil pengamatan di beberapa perkebunan karet menunjukkan bahwa sekitar 22,2% mutu sadap DC tergolong baik, sedangkan 44,4% tergolong cukup baik, dan 33,3% tergolong jelek.
Sistem sadap DC sudah memiliki panduan agar perolehan produksinya optimal, yaitu tergambar dalam tataguna panel sebagai berikut:
Idealnya, penyadapan DC dapat berlangsung selama 8 tahun, yaitu sejak TM ke-11 s.d ke-18. Potensi perolehan produktivitas per hektar per tahun mencapai sekitar 2 ton/ha/th. Mutu sadap yang jelek dan kontrol sadap yang lemah mengakibatkan potensi tersebut tidak dapat dicapai. GPS atau produktivitas individu tanaman (gr/pohon/sadap) pada kondisi demikian biasanya juga sangat rendah, hanya berkisar 30-35 gr/pohon/sadap atau bahkan kurang. Padahal potensi atau idealnya adalah sekitar 50 gr/pohon/sadap.
Potensi produksi dan umur ekonomi tersebut dapat dicapai jika penyadapan DC dilakukan secara benar, yaitu sebagai berikut:
- Apakah semua pohon tuntas disadap?
- Apakah semua panel tuntas disadap (jangan ada sadap pincang)?
- Apakah stimulan sudah efektif?
- Apakah waktu / durasi menyadap normal? dsb
DAFTAR PUSTAKA
Akhmad Rouf, Mudita Oktorina Nugrahani, dan Yoga Bagus Setya Aji. 2020. Penerapan Sistem Sadap Irisan Ganda (Double Cut) Pada Klon Karet Slow Starter (Studi kasus di beberapa perkebunan karet wilayah Jawa). Pros. Semnas PKM Center UNS. Hal: 194-198.
Sumarmadji, Akhmad Rouf, Yoga Bagus Setya Aji, dan Titik Widyasari. 2017. Optimalisasi Produksi dan Penekanan Biaya Penyadapan dengan Sistem Sadap Intensitas Rendah. Warta Perkaretan 2017 Vol 36 No 1. Hal: 39-54.
Sumarmadji, Junaidi, Atminingsih, Kuswanhadi, Akhmad Rouf. 2012. Paket teknologi penyadapan untuk optimasi produksi sesuai tipologi klon. Pros. Konf. Nas. Karet. Yogyakarta, 19-20 September 2012. Pusat Penelitian Karet. Hal: 207-216.
(Akhmad Rouf – Kelti Agronomi / Fisiologi Penyadapan Tanaman Karet)

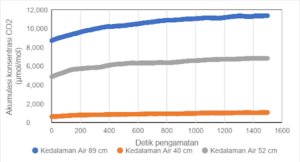
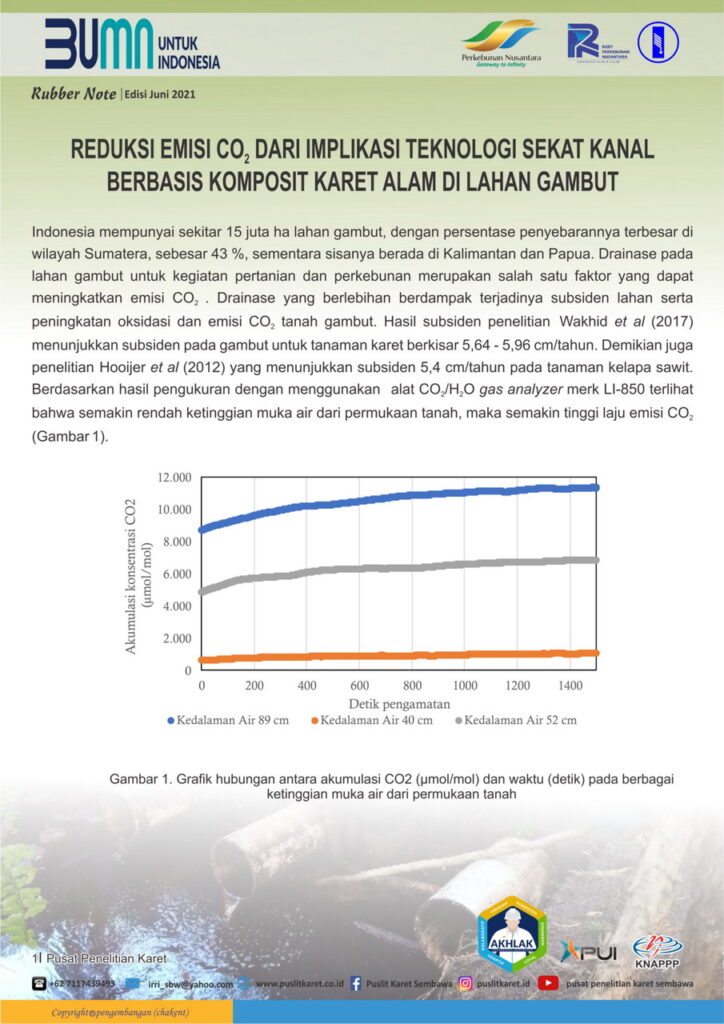
Indonesia mempunyai sekitar 15 juta ha lahan gambut, dengan persentase penyebarannya terbesar di wilayah Sumatera, sebesar 43 %, sementara sisanya berada di Kalimantan dan Papua. Drainase pada lahan gambut untuk kegiatan pertanian dan perkebunan merupakan salah satu faktor yang dapat meningkatkan emisi CO2 . Drainase yang berlebihan berdampak terjadinya subsiden lahan serta peningkatan oksidasi dan emisi CO2 tanah gambut. Hasil subsiden penelitian Wakhid et al (2017) menunjukkan subsiden pada gambut untuk tanaman karet berkisar 5,64 - 5,96 cm/tahun. Demikian juga penelitian Hooijer et al (2012) yang menunjukkan subsiden 5,4 cm/tahun pada tanaman kelapa sawit. Berdasarkan hasil pengukuran dengan menggunakan alat CO2/H2O gas analyzer merk LI-850 terlihat bahwa semakin rendah ketinggian muka air dari permukaan tanah, maka semakin tinggi laju emisi CO2 (Gambar 1).
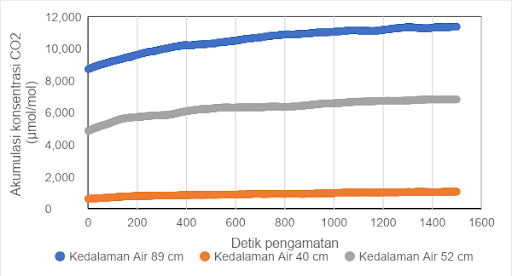
Gambar 1. Grafik hubungan antara akumulasi CO2 (µmol/mol) dan waktu (detik) pada berbagai ketinggian muka air dari permukaan tanah
Tata kelola air merupakan kunci untuk mengurangi emisi CO2. Pengaturan tinggi muka air tanah di lahan gambut ini dapat dilakukan dengan teknologi sekat kanal. Saat ini, sebagian besar pembuatan kanal yang dilakukan masyarakat maupun perusahaan belum mempertimbangkan pengaturan muka air dari lahan gambut. Kelemahan pembangunan sekat kanal yang dibuat secara tradisional dengan desain sederhana menggunakan kayu dan karung pasir adalah mudah bocor sehingga ketinggian muka air tidak dapat terjaga. Salah satu upaya untuk mengatasi hal tersebut adalah melalui teknologi sekat kanal berbasis komposit karet alam untuk mengatur drainase. Komposit karet alam memiliki kelebihan kedap air dan material penyokong sekat kanal seperti pasir, split, dan kerikil tidak mudah terhanyut.
Hasil implementasi dari teknologi sekat kanal berbasis komposit karet alam terbukti mampu menjaga tinggi muka air saat musim kering dan menurunkan subsiden pada lahan gambut. Selama periode musim kering (curah hujan < 50 mm), sekat kanal berbasis komposit karet alam mampu menahan air sebesar ± 15 cm di lahan gambut (Gambar 1). Reduksi emisi CO2 melalui perbedaan ketinggian muka air berdasarkan metode empirik adalah 34,73 ton CO2-eq/ha/hari atau setara 12.506 ton CO2-eq/ha/tahun.
 |
 |
|
Gambar 1. Perbedaan ketinggian muka air didalam (A) dan diluar sekat kanal (B) saat periode musim kering. Perbedaan ketinggian muka air ditnadai dengan air yang mengalir dari dalam ke luar sekat kanal |
|
Daftar Pustaka :
Hooijer, A., Page, S., Jauhiainen, J., Lee, W. A., Lu, X. X., Idris, A., & Anshari, G. (2012). Subsidence and carbon loss in drained tropical peatlands. Biogeosciences, 9(3), 1053–1071. https://doi.org/10.5194/bg-9-1053-2012
Wakhid, N., Hirano, T., Okimoto, Y., Nurzakiah, S., & Nursyamsi, D. (2017). Soil carbon dioxide emissions from a rubber plantation on tropical peat. Science of The Total Environment, 581–582, 857–865. https://doi.org/10.1016/j.scitotenv.2017.01.035